

A Case Study of Bioenergy Crops: Increasing the utility of "Populus"



 To produce second-generation biofuels, cellulosic feedstocks will be needed from a variety of sources. Using a single gene, researchers from Purdue University in Indiana have found that it’s possible to affect many of the traits in poplar (a species within the genus Populus) that are desirable for energy crops. This approach could be advantageous not only for poplar, but also for other crops that have potential as cellulosic feedstocks, possibly heightening production and lowering the cost of biofuels.
To produce second-generation biofuels, cellulosic feedstocks will be needed from a variety of sources. Using a single gene, researchers from Purdue University in Indiana have found that it’s possible to affect many of the traits in poplar (a species within the genus Populus) that are desirable for energy crops. This approach could be advantageous not only for poplar, but also for other crops that have potential as cellulosic feedstocks, possibly heightening production and lowering the cost of biofuels.
Some traits desired for most, if not all, dedicated energy crops include:
1. Vegetative propagation, for asexual production of rooted plants from cuttings;
2. Rapid growth;
3. High conversion efficiency;
4. Altered lignin composition and/or content;
5. Reproductive sterility, to avoid diversion of photosynthate away from vegetative growth and for transgene confinement; and
6. Growth distributed across several stems, rather than one central stem, for ease of harvesting using existing farm equipment.
Research for growth
In plants, a class of small RNA molecules, called microRNAs, function as regulators of various developmental events. They perform this role by binding to the messenger RNA (mRNA) of genes they target for degradation.
For the purposes of this case study, the Corngrass1 (Cg1) gene was isolated from a mutant maize (Zea mays) plant in which it was over-expressed. This mutant plant exhibited the following traits: adventitious root formation, enabling vegetative propagation; pronounced shoot proliferation; and partial sterility, due to alterations in meristem development.
Tobacco (Nicotiana tabacum) and Arabidopsis thaliana plants that were genetically engineered (GE) to express Cg1 constitutively (i.e., at high levels, in all tissues, all the time), and produced multiple axillary shoots, grew faster than the wild-type plants, contained less lignin, and were either sterile or exhibited delayed flowering.
These findings with herbaceous plants led the Purdue researchers to investigate the effect of expressing Cg1 in poplar, which are trees that are capable of rapid growth and that can be propagated asexually. The phenotypes seen in the GE poplar are consistent with those seen other heterologous species in which Cg1 has been over-expression, including altered lignin content.
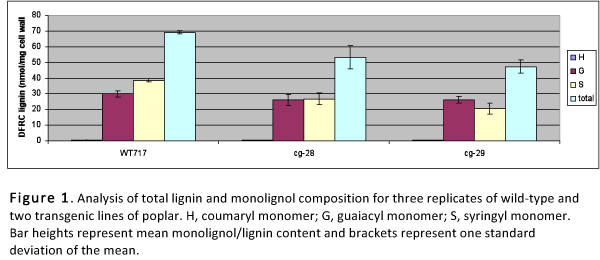
In addition, changes in lignin composition were seen. Most plants, including poplar, use syringyl (S) and guaiacyl (G) monomers to synthesize lignin, an important cell-wall polymer. The ratio of S to G units is fixed for a given species, and affects the strength with which lignin is bound to other components of the plant cell wall. The GE poplars both had altered S:G ratios and reduced lignin content, which makes it easier to extract, thereby improving conversion efficiency.
In summary, using a single regulatory gene from maize, it’s possible to induce in poplar all of the traits desired for bioenergy crops. Low conversion efficiency is a problem for virtually all cellulosic feedstocks, and not just poplar. Therefore, results seen with poplar may be used to inform efforts being made to improve other bioenergy crops, as well.
Flowering control
The only phenotype that hasn’t been verified in the GE poplar is flowering control. This is their second growing season in the field, under a permit issued by Biotechnology Regulatory Services unit of the Animal Plant Health Inspection Service (APHIS), but poplars have a juvenile period of three to five years. So, it can’t be determined whether the introduced Cg1 gene will impart sterility until the trees are competent to produce flowers.
Currently, transgenic trees can only be grown in the field for research purposes, and only for three years. With suitable justification, it’s possible to obtain an extension. Federal regulators are likely to approve a request to allow these GE poplars to remain in the field beyond three years for a few reasons. First, these transgenic trees are female (poplars are dioecious) and, therefore, don’t produce pollen, only seed. Second, vegetative and reproductive buds are morphological distinct, so the latter could be removed before flowers emerge in the spring. Finally, there are not sexually compatible, wild relatives in the growing in the vicinity of the field trial.
Because trees have not been domesticated to the same extent as annual, agronomic crops, they have wild relatives with which they are inter-fertile. All cells produced by GE plants contain the introduced gene (“transgene”), including the cells that make up their pollen. Many trees, including poplar, are obligate out-crossers and wind pollinated. Federal regulators are concerned that if a GE tree is allowed to flower, its pollen will fertilize non-transgenic trees outside a plantation, in the wild. Seed produced as a result of fertilization by pollen from a GE tree will contain the transgene, but there will be no way of knowing which seedlings derived from this seed contain the transgene based on their physical appearance.
Depending on the trait it controls, the transgene could affect the invasiveness or weediness of plants possessing it. Until a way can be found to prevent the spread of transgenes in the environment, federal regulators are very unlikely to allow GE trees to be grown for commercial purposes. Reproductive sterility is one way of achieving this transgene confinement. If the poplars expressing Cg1 don’t flower, they have the potential to be deployed directly, and they would be commercially desirable because of their rapid growth (photosynthate won’t be diverted away from vegetative growth for the development of reproductive structures, such as flowers, seeds, and cones).
The future
The research with Cg1 has even greater potential. Corngrass1 controls the expression of other genes by way of a very specific targeting site. More than a dozen genes in the poplar genome contain this highly conserved sequence. None of the candidate poplar genes have been annotated. In other words, although it’s known that these sequences encode genes, the functions performed by these targeted genes are unknown.
By determining the roles of these target genes, it could be possible to tailor trees for specific end uses (e.g., direct combustion, pyrolysis for producing a “green diesel,” fermenting the sugars contained in the feedstock to produce ethanol, etc.), rather than altering the entire suite of genes simultaneously.
Rick Meilan is an Associate Professor of Molecular Tree Physiology in the Department of Forestry and Natural Resources at Purdue University.
Purdue University
www.purdue.edu
www.agriculture.purdue.edu/fnr/html/faculty/Meilan/index.htm
Author: Rick Meilan
Volume: November/December 2012




